
NK-клетки относятся к врожденному звену иммунитета, им не требуется предварительной сенсибилизации для осуществления защитных функций. При беременности NK-клетки мигрируют в ткань матки, где участвуют в регуляции формирования и развития ткани плаценты. € ‚ƒ„…
…Особенности функциональных характеристик NK-клеток
Морфологически NK-клетки представляют собой большие гранулярные CD3-негативные лимфоциты, экспрессирующие поверхностную молекулу CD56. В настоящее время по характеру экспрессии поверхностных маркеров NK-клетки можно подразделить на 48 различных популяций. Однако традиционно среди NK-клеток выделяют две группы: CD3– CD16bright CD56dim и CD3– CD16dim CD56bright. Большинство NK-клеток периферической крови слабо экспрессируют CD56 и интенсивно экспрессируют CD16 (CD16bright CD56dim). Для этих клеток характерно наличие большого количества лизосомальных гранул, содержащих гранзимы и перфорины. Другие популяции NK-клеток периферической крови экспрессируют CD56, но слабо или совсем не экспрессируют молекулы CD16 (CD16dim CD56bright и CD16–CD56bright). Популяции CD16dim CD56bright и CD16–CD56bright обладают регуляторными свойствами, однако под воздействием цитокинов свойства и функции этих NK-клеток могут изменяться. Так, интерлейкин (IL)-2 модулирует как фенотип, так и цитотоксическую функцию
NK-клеток: в экспериментах in vitro при стимуляции IL-2 CD16dim CD56bright NK-клеток уровень цитотоксичности этой популяции NK-клеток становился сходным с уровнем цитотоксичности СD16bright CD56dim NK-клеток.
Дифференцировка разных популяций NK-клеток происходит из общего предшественника лимфоидных клеток под воздействием цитокина IL-15, продуцируемого стромальными клетками костного мозга [2]. На начальных этапах дифференцировки гемопоэтические стволовые клетки приобретают фенотип CD34bright CD122+ (IL-2Rβ+ IL-15Rβ+) CD56– и в ответ на стимуляцию IL-15 развиваются в зрелые NK-клетки. Такой путь дифференцировки характерен для преобладающих в периферической крови CD16bright CD56dim NK-клеток. В лимфатических узлах основной пул стволовых клеток составляют клетки-предшественники с фенотипом CD34dim, конститутивно экспрессирующие CD45RA,
L-селектин, LFA-1 (lymphocyte function-associated antigen-1), β7-интегрин и высокоаффинный рецептор к IL-2 – IL-2Rαβγ. Эти гемопоэтические стволовые клетки, мигрировавшие в лимфатические узлы из костного мозга, могут дифференцироваться в CD16dim CD56bright NK-клетки после стимуляции IL-2 или IL-15 [14]. При этом клетки с фенотипом CD34dim CD45RA+β7high не являются предшественниками CD56dim NK-клеток, что дает основание говорить о двух независимых путях дифференцировки NK-клеток. Однако после стимуляции IL-2 CD16dim CD56bright NK-клетки лимфатических узлов могут приобретать фенотип CD16bright CD56dim и проявлять цитотоксические свойства [12]. По другим
данным, CD16bright CD56dim NK-клетки в ткани при воздействии на них IL-12 могут изменять свой
фенотип на CD16dim CD56bright [24]. Таким образом, дифференцировка основной популяции NK-клеток периферической крови (CD16bright CD56dim) происходит в костном мозге, в дальнейшем под воздействием IL-12 в ткани они могут приобретать фенотип CD16dim CD56bright. Дифференцировка CD16dim CD56bright NK-клеток также происходит в костном мозге, однако более точный фенотип предшественников этой популяции NK-клеток еще предстоит уточнить. Основным же путем образования CD16dim CD56bright NK-клеток является их дифференцировка из клеток-предшественников в лимфатических узлах (рисунок, см. на вклейке).
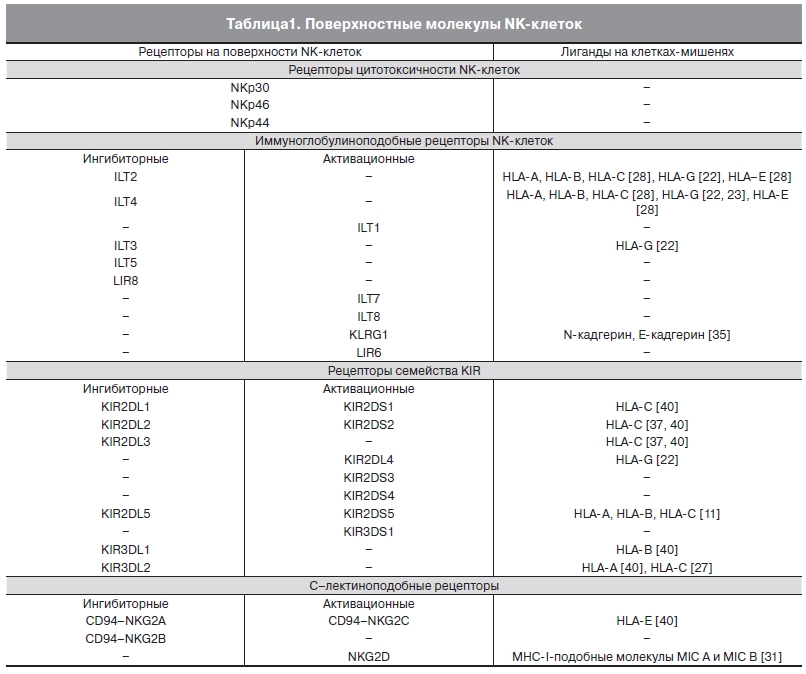
При связывании NK-клеток с клеткой-мишенью образуется иммунологический синапс. В случае если клетка-мишень является вирусинфицированной или опухолевой, формируется межклеточный комплекс, передающий активационный сигнал NK-клетке. Формирование иммунологического синапса и запуск дальнейших реакций представляют собой процесс, включающий взаимодействия адгезионных молекул [21], С-лектиноподобных рецепторов, рецепторов KIR NK-клеток [31] с лигандами на клетках-мишенях. Контакт NK-клетки с клеткой-мишенью вызывает агрегацию фибриллярного актина, талина и других белков цитоскелета [21], что приводит к высвобождению перфоринов и гранзимов. При контакте NK-клетки с клеткой-мишенью, несущей на своей мембране MHC-I (major histocompatibility complex) с собственным клеточным пептидом, формируется ингибиторный иммунологический синапс, препятствующий активации NK-клетки. Экспрессия большинства MHC-I специфичных рецепторов на NK-клетках клоноспецифична: для NK-клеток характерна экспрессия как минимум одного ингибирующего рецептора, при этом активационные рецепторы NK-клетка может не экспрессировать [5]. В табл. 1 приведены данные о рецепторах NK-клеток и их лигандах на мембране клеток-мишеней. Следует отметить, что в настоящее время значительная часть лигандов рецепторов NK-клеток не выявлена.
Таблица 1. Поверхностные молекулы NK-клеток. „
Роль NK-клеток при физиологическом течении беременности ‚†‡ €‡
NK-клетки, присутствующие в эндометрии матки, играют важную роль в процессе имплантации и в
дальнейшем развитии беременности. В течение пролиферативной фазы менструального цикла NK-клетки матки представляют собой небольшую популяцию клеток. Однако их количество в эндометрии значительно возрастает в секреторную фазу цикла [34]. После имплантации зародыша в ходе децидуализации эндометрий матки претерпевает значительные изменения, одно из которых –
появление большого количества децидуальных NK-клеток с фенотипом CD3—CD56bright, составляющих до 70% всех лимфоцитов, присутствующих в децидуальной ткани, и примерно 30% всех клеток децидуальной ткани [2]. Для NK-клеток матки характерна высокая экспрессия CD56 и низкая экспрессия CD16. Лишь небольшое количество NK-клеток являются CD56dim [34]. По некоторым данным, CD3—CD56bright NK-клетки присутствуют в большом количестве в слизистой оболочке матки
еще до имплантации. В случае наступления беременности NK-клетки матки аккумулируются вокруг
клеток трофобласта, а максимальное их содержание совпадает с периодом инвазии трофобласта [5].
В настоящее время дискуссионным остается вопрос о процессе образования и пополнения пула NK-клеток матки при беременности. CD16dim CD56bright NK-клетки периферической крови и децидуальной ткани экспрессируют хемокиновый рецептор CXCR3 (рецептор к CXCL10: IP-10 – γ-интерферониндуцибельному протеину) и CXCR4, лигандом которого является SDF-1 (Stromal cell-derived factor-1) (CXCL12) [15, 39]. При этом для CD16dim CD56bright NK-клеток при беременности характерно увеличение как количества клеток, экпрессирующих CXCR4, так и интенсивности его экспрессии на этих клетках. Для децидуальной ткани в отличие от стромы ворсин хориона характерна экспрессия хемокинов CXCL12 (лиганд CXCR4) и CXCL10 (лиганд CXCR3). Кроме того, клетки экстравиллезного трофобласта также экспрессируют CXCL12 [15, 39]. In vitro клетки стромы матки и децидуальные эндотелиальные клетки в I тримеcтре беременности экспрессируют мРНК CXCL10 (IP-10). В экспериментах in vitro показано, что CD16dim CD56bright NK-клетки периферической крови при стимуляции CXCL10 (IP-10) мигрируют через монослой децидуальных эндотелиальных клеток и клетки стромы матки [8]. При культивировании CD16bright CD56dim NK-клеток в присутствии супернатантов, полученных после культивирования стромальных клеток матки, наблюдается снижение экспрессии NK-клетками CD16 и изменение их фенотипа с CD16bright на CD16dim. Смену фенотипа NK-клеток вызывает цитокин трансформирующий фактор роста β (TGF-β –Transforming growth factor), продуцируемый децидуальными клетками. Также в децидуальной ткани обнаружены гемопоэтические стволовые клетки, которые при культивировании в присутствии IL-15 и фактора стволовых клеток SCF (stem cell factor) приобретают фенотип CD16–CD56bright децидуальных NK-клеток [18].
Ранее установлено, что NK-клетки экспрессируют такие адгезионные молекулы, как LFA-1 [6], Mac-1 [21], PECAM-1 (platelet-endothelial cell adhesion molecule 1) [38]. Лиганды к адгезионным молекулам NK-клеток выявлены на эндотелиальных клетках и клетках трофобласта (табл. 2).
Таблица 2. Экспрессия адгезионных молекул NK-клетками и их лигандов на клетках эндотелия и трофобласта.
Экспрессия молекул адгезии опосредует как миграцию NK-клеток в децидуальную ткань из периферической крови, так и взаимодействие NK-клеток с клетками трофобласта. Таким образом, пул
NK-клеток матки формируется в начале I триместра беременности за счет миграции NK-клеток
из периферической крови. Клетки стромы матки, эндотелиальные клетки децидуальной оболочки
и клетки трофобласта за счет секреции хемокинов контролируют хемотаксис NK-клеток с фенотипом CD16dim CD56bright из периферической крови. Пополнение пула NK-клеток матки может происходить за счет CD16bright CD56dim NK-клеток периферической крови. Под воздействием цитокинов CD16bright CD56dim NK-клетки изменяют свой фенотип на CD16dim CD56bright и приобретают свойства, характерные для CD16dim CD56bright NK-клеток. Затем увеличение количества CD16dim CD56bright
NK-клеток в децидуальной ткани может происходить за счет пролиферации in situ.
Клетки плода несут антигены как матери, так и отца, что может вызывать активацию NK-клеток матки. Возникновение толерантности иммунной системы матери к антигенам плода объясняют взаимодействием рецепторов децидуальных лимфоцитов с молекулами локуса HLA-G, экспрессированных на клетках трофобласта. Лигандами для молекул локуса HLA-G являются рецепторы ILT и KIR2DL4 NK-клеток. Взаимодействие этих рецепторов с молекулами локуса HLA(human leukocyte antigen) –G препятствует активации цитотоксической функции NK-клеток. Однако блокирование рецептора KIR2DL4 NK-клеток с помощью специфичных антител не приводит к проявлению цитотоксических свойств децидуальных NK-клеток по отношению к клеткам трофобласта [5]. Следовательно, лиганд-рецепторные взаимодействия клеток трофобласта и NK-клеток не сводятся исключительно к ингибированию цитотоксичности последних.
Имплантация и нормальное развитие беременности во многом зависят и от продуцируемых децидуальными NK-клетками цитокинов. Беременность сопровождается повышенной секрецией интерферона-γ (IFN-γ) NK-клетками. Связывание молекулы локуса HLA-G, экспрессированной на клетках трофобласта, и рецептора KIR2DL4 NK-клеток стимулирует секрецию IFN-γ NK-клетками при одновременном ингибировании их цитотоксической активности в отношении клеток трофобласта [36]. В экспериментах in vivo показано, что при взаимодействии рецептора NKG2D NK-клеток с лигандом RAET1 (retinoic acid early transcript 1) клеток трофобласта также индуцируется секреция IFN-γ NK-клетками [7]. IFN-γ способствует формированию ткани плаценты, снижая миграцию клеток трофобласта и обеспечивая их скопление вблизи спиральных артерий [17]. При воздействии на эндотелиальные клетки IFN-γ индуцирует экспрессию и секрецию молекул локуса HLA-E эндотелиальными клетками, что препятствует их лизису NK-клетками [9].
Неотъемлемой частью физиологического развития беременности является апоптоз клеток трофобласта при формировании и перестройках ткани плаценты. Запуск процесса апоптоза индуцируется лигандрецепторными взаимодействиями TRAIL/TRAIL-R и Fas/FasL. Для клеток трофобласта характерна экспрессия Fas, FasL и рецепторов TRAIL – TRAIL-R1 и TRAIL-R2. NK-клетки матки экспрессируют FasL, при связывании которого с Fas происходит индукция их гибели. Также для NK-клеток характерна экспрессия TRAIL, который при связывании с рецептором может индуцировать гибель клеток трофобласта. В целом апоптоз клеток децидуальной оболочки и плаценты в норме при беременности носит ограниченный характер: большинство клеток трофобласта не подвергается апоптозу за счет ингибиторов апоптоза – FLIP (fice-like inhibitory protein) и XIAP (X-linked inhibitor of apoptosis), препятствующих проведению внутриклеточного сигнала от Fas [33]. Кроме того, под воздействием IL-15, цитокина, присутствующего в ткани плаценты при физиологическом течении беременности, NK-клетки экспрессируют рецепторы TRAIL-R2 и TRAIL-R3 [26] и, таким образом, могут сами подвергаться апоптозу в случае активации и угрозы нормальному развитию беременности.
NK-клетки матки оказывают иммуномодулирующее действие на клетки плаценты за счет продукции гликоделина и галектина-1. Гликоделин и галектин-1 обладают подавляющими иммунный ответ свойствами и влияют на цитокиновую сеть плаценты. Они уменьшают продукцию фактора некроза
опухоли α (TNF-α – tumor necrosis factor), IL-2 и IFN-γ Т-лимфоцитами, локализованными в эндометрии, а также снижают продукцию IL-12 активированными макрофагами, что способствует индукции иммунной толерантности в отношении плода.
Для децидуальных NK-клеток показана высокая экспрессия MIP-1a, GM-CSF, колониестимулирующего фактора-1 по сравнению с CD16dim CD56bright NK-клетками периферической крови. Также NK-клетки матки продуцируют ангиопоэтин-2 и плацентарный фактор роста (PlGF – placental growth factor) [24]. В экспериментах in vitro при стимуляции NK-клеток антителами к рецепторам KIR2DL4 в течение 2 ч после стимуляции возрастает транскрипция генов IL-1β, IL-8, MIP-3α, а затем генов IL-6, IL-12β и IL-23α [30]. При физиологической беременности в децидуальной ткани присутствуют растворимые молекулы HLA-G трофобласта. При связывании с ними рецепторов KIR2DL4 NK-клетки экспрессируют IL-1β и IFN-γ, стимулируют продукцию фактора роста эндотелия сосудов (VEGF – Vascular endothelial growth factor) клетками трофобласта и таким образом способствуют ангиогенезу и повышают жизнеспособность клеток плаценты [30]. Кроме того, NK-клетки продуцируют IL-8 и IP-10, цитокины, способствующие инвазии трофобласта [20]. Таким образом, NK-клетки матки оказывают значительное влияние на процессы формирования ткани плаценты. Изменение функционального состояния NK-клеток может приводить к нарушению формирования ткани плаценты и развитию таких патологий беременности, как невынашивание беременности и гестоз.
Гестоз – одно из наиболее опасных осложнений беременности, эта патология развивается после
20-й недели беременности и является причиной заболеваемости и смертности беременных женщин
[1]. В настоящее время гестоз рассматривают как полиэтиологичное заболевание, характеризующееся нарушением иммунной адаптации в системе мать–плод. Одной из причин нарушения толерантности иммунной системы матери к плоду при гестозе является недостаточная экспрессия молекулы локуса HLA-G на клетках трофобласта. В этом случае не происходит связывания молекулы HLA-G с рецептором KIR2DL4 на NK-клетках, цитотоксическая активность децидуальных NK-клеток не подавляется и вследствие этого развивается местная воспалительная реакция [1]. В исследовании фенотипа NK-клеток матки у беременных с гестозом установлено, что количество CD3—CD16+ NK-клеток возрастает в периферической крови при гестозе по сравнению с таковым при физиологическом течении беременности [4, 5]. В ткани плаценты при гестозе также значительно увеличивается количество продуцируемого NK-клетками IFN-γ при одновременном снижении продукции IL-10 клетками плаценты [3]. Это приводит к преобладанию провоспалительных цитокинов в плаценте и активации клеток иммунной системы матери в отношении антигенов плода и развитию гестоза. В норме индукции иммунной толерантности при беременности способствует экспрессируемая дендритными клетками и макрофагами плаценты индоламин-2,3-диоксигеназа. Этот фермент препятствует активации Т-лимфоцитов и NK-клеток. При гестозе наблюдается снижение продукции этого фермента [32], что способствует активации NK-клеток матери и проявлению цитотоксической активности.
При гестозе наблюдается повышенная экспрессия FasL децидуальными лейкоцитами и, в частности, NK-клетками [25]. Связывание рецепторов Fas клеток трофобласта и FasL NK-клеток приводит к усиленной гибели клеток трофобласта, что в итоге приводит к нарушению формирования ткани пла-
центы. При физиологическом течении беременности индукции апоптоза клеток трофобласта препятствуют экспрессируемые трофобластом ингибиторы апоптоза IAP (inhibitors of apoptosis). В контроле их экспрессии участвуют белки митохондрий, например smac. При гестозе выявлено снижение экспрессии этого белка в клетках трофобласта [16], что свидетельствует о невозможности клеток трофобласта компенсировать апоптогенную активность NK-клеток. Беременность, осложненная гестозом, также сопровождается снижением концентрации IL-15 в ткани плаценты, что способствует
снижению экспрессии NK-клетками рецепторов TRAIL-R2 и TRAIL-R3 и также приводит к нарушению механизма защиты клеток трофобласта от цитотоксичеких эффектов NK-клеток.
Другой распространенной патологией беременности является привычное невынашивание. К развитию этой патологии могут приводить внутриутробная инфекция, нарушения эндокринной регуляции, анатомические особенности матки, нарушение иммунной толерантности в системе мать – плод. При этой патологии наблюдается достоверное увеличение количества CD16bright CD56dim NK-клеток в периферической крови [19] и в ткани плаценты [10], а также уменьшение количества CD16dim CD56bright NK-клеток в периферической крови [19]. Увеличение количества цитотоксических NK-клеток CD16bright CD56dim может способствовать индукции цитотоксических реакций в ткани плаценты. Так, для NK-клеток при невынашивании показано уменьшение экспрессии ингибиторных рецепторов KIR (KIR2DL2) [13]. При этом сниженная экспрессия молекул локуса HLA-G клетками плаценты, как и уменьшение экспрессии ингибиторных рецепторов NK-клетками, приводит к проявлению цитотоксических свойств децидуальных NK-клеток. Резкое увеличение количества NK-клеток в ткани матки на начальных стадиях развития беременности приводит к интенсификации ангиогенеза и нарушению формирования ткани плаценты, резкому повышению концентрации кислородных радикалов и как следствие невынашиванию беременности [29].
Суммируя приведенные выше данные литературы, необходимо отметить, что при физиологическом течении беременности NK-клетки мигрируют из периферической крови в ткань матки, где происходит изменение их фенотипа с цитотоксического (CD16bright CD56dim) на регуляторный (CD16dim CD56bright). При этом NK-клетки матки обладают сниженной цитотоксической активностью при сохранении способности к синтезу и секреции IFN-γ и других цитокинов. При физиологическом течении беременности NK-клетки остаются иммунологически толерантными в отношении клеток трофобласта и плаценты. При нарушении экспрессии клетками трофобласта неклассических молекул главного комплекса гистосовместимости HLA-G цитотоксическая активность NK-клеток матки не подавляется, что способствует индукции местной воспалительной реакции в ткани плаценты, нарушению формирования сосудистой сети плаценты за счет усиления апоптотической гибели клеток плаценты. Нарушение экспрессии NK-клетками рецепторов KIR, взаимодействующих с неклассическими молекулами локуса HLA I класса, при беременности приводит к усилению цитотоксической активности NK-клеток, что также способствует развитию таких осложнений беременности, как гестоз и привычное невынашивание.



{kind=link}
{kind=link}